Медичний експерт статті

Нові публікації
Яєчник
Останній перегляд: 04.07.2025

Весь контент iLive перевіряється медичними експертами, щоб забезпечити максимально можливу точність і відповідність фактам.
У нас є строгі правила щодо вибору джерел інформації та ми посилаємося тільки на авторитетні сайти, академічні дослідницькі інститути і, по можливості, доведені медичні дослідження. Зверніть увагу, що цифри в дужках ([1], [2] і т. д.) є інтерактивними посиланнями на такі дослідження.
Якщо ви вважаєте, що який-небудь з наших матеріалів є неточним, застарілим або іншим чином сумнівним, виберіть його і натисніть Ctrl + Enter.
Яєчник (оваріум; грец. оопхорон) — парний орган, жіноча статева залоза, розташована в порожнині малого таза позаду широкої зв'язки матки. У яєчниках розвиваються та дозрівають жіночі статеві клітини (яйцеклітини), утворюються жіночі статеві гормони, що потрапляють у кров і лімфу. Яєчник має яйцеподібну форму, дещо сплющену в передньозадньому напрямку. Колір яєчника рожевий. На поверхні яєчника жінки, яка народила, видно заглиблення та рубці — сліди овуляції та перетворення жовтого тіла. Маса яєчника становить 5-8 г. Розміри яєчника: довжина 2,5-5,5 см, ширина 1,5-3,0 см, товщина — до 2 см. Яєчник має дві вільні поверхні: медіальну поверхню (facies medialis), звернену до порожнини малого таза, частково покриту матковою трубою, та латеральну поверхню (facies lateralis), що прилягає до бічної стінки таза, до слабо вираженого заглиблення — яєчникової ямки. Ця ямка розташована в куті між зовнішніми клубовими судинами, покритими очеревиною зверху, та матковою та запиральною артеріями знизу. Позаду яєчника ретроперитонеально зверху вниз проходить сечовід відповідної сторони.
Поверхні яєчника переходять у опуклий вільний (задній) край (margo liber), спереду – у брижовий край (margo mesovaricus), прикріплений за допомогою короткої складки очеревини (брижі яєчника) до заднього листка широкої зв'язки матки. На цьому передньому краї органу є жолобчасте заглиблення – ворота яєчника (hilum ovarii), через яке в яєчник входять артерія та нерви, а виходять вени та лімфатичні судини. Яєчник також має два кінці: закруглений верхній трубчастий кінець (extremitas tubaria), звернений до маткової труби, та нижній матковий кінець (extremitas utenna), з'єднаний з маткою зв'язкою яєчника (lig. ovarii proprium). Ця зв'язка, у вигляді круглого тяжа товщиною близько 6 мм, йде від маткового кінця яєчника до латерального кута матки, розташованого між двома листками широкої зв'язки матки. До зв'язкового апарату яєчника також належить зв'язка, що підвішує яєчник (lig.suspensorium ovarii), що являє собою складку очеревини, що проходить від стінки малого таза до яєчника та містить усередині судини яєчника та пучки фіброзних волокон. Яєчник фіксується короткою брижею (mesovarium), що являє собою дуплікацію очеревини, що проходить від заднього листка широкої зв'язки матки до брижового краю яєчника. Самі яєчники очеревиною не покриті. Найбільший яєчниковий краєчок маткової труби прикріплюється до трубчастого кінця яєчника. Топографія яєчника залежить від положення матки, її розмірів (під час вагітності). Яєчники є дуже рухливими органами порожнини малого тазу.
 [
[ Судини та нерви яєчника
Кровопостачання яєчників забезпечується aa. et vv. ovaricae et uterinae. Обидві яєчникові артерії (aa. ovaricae dextra et sinistra) беруть початок від передньої поверхні аорти трохи нижче ниркових артерій; права зазвичай бере початок від аорти, а ліва - від ниркової артерії. Прямуючи вниз і латерально вздовж передньої поверхні великого поперекового м'яза, кожна яєчникова артерія перетинає спереду сечовід (віддаючи до нього гілки), зовнішні клубові судини, прикордонну лінію та входить у порожнину малого таза, розташовуючись тут у підвішувальній зв'язці яєчника. Слідуючи в медіальному напрямку, яєчникова артерія проходить між листками широкої зв'язки матки під матковою трубою, віддаючи до неї гілки, а потім у брижу яєчника; вона входить у ворота яєчника.
Гілки яєчникової артерії широко анастомозують з яєчниковими гілками маткової артерії. Венозний відтік від яєчників здійснюється переважно в яєчникове венозне сплетення, розташоване в області воріт яєчника. Звідси відтік крові проходить у двох напрямках: через маткові та яєчникові вени. Права яєчникова вена має клапани та впадає в нижню порожнисту вену. Ліва яєчникова вена впадає в ліву ниркову вену, хоча клапанів не має.
Лімфатичний відтік від яєчників відбувається по лімфатичних судинах, особливо рясно в ділянці воріт органу, де виділяється субоваріальне лімфатичне сплетення. Потім лімфа відводиться по лімфатичних судинах яєчників до парааортальних лімфатичних вузлів.
Іннервація яєчників
Симпатична – забезпечується постгангліонарними волокнами від черевного (сонячного), верхнього брижового та гіпогастрального сплетень; парасимпатична – вісцеральними крижовими нервами.
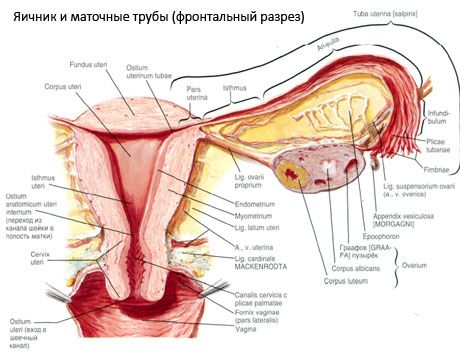
Будова яєчника
Поверхня яєчника покрита одношаровим зародковим епітелієм. Під ним лежить щільна сполучнотканинна білкова оболонка (tunica albuginea). Сполучна тканина яєчника утворює його строму (stroma ovarii), багату на еластичні волокна. Речовина яєчника, його паренхіма, поділяється на зовнішній та внутрішній шари. Внутрішній шар, розташований у центрі яєчника, ближче до його воріт, називається мозковою речовиною (medulla ovarii). У цьому шарі, в пухкій сполучній тканині, знаходяться численні кровоносні та лімфатичні судини та нерви. Зовнішній шар яєчника - кіркова речовина (cortex ovarii) є щільнішим. Вона містить багато сполучної тканини, в якій розташовані дозріваючі первинні фолікули яєчника (folliculi ovarici primarii), вторинні (везикулярні) фолікули (folliculi ovarici secundarii, s.vesiculosi), а також зрілі фолікули, граафові фолікули (folliculi ovarici maturis), а також жовті та атретичні тільця.
Кожен фолікул містить жіночу статеву яйцеклітину, або ооцит (овоцит). Яйцеклітина має діаметр до 150 мкм, круглу форму, містить ядро, велику кількість цитоплазми, яка, крім клітинних органел, містить білково-ліпідні включення (жовток), глікоген, необхідні для живлення яйцеклітини. Яйцеклітина зазвичай витрачає свій запас поживних речовин протягом 12-24 годин після овуляції. Якщо запліднення не відбувається, яйцеклітина гине.
Людська яйцеклітина має дві мембрани, що покривають її. Всередині знаходиться цитолема, яка є цитоплазматичною мембраною яйцеклітини. Зовні цитолемми знаходиться шар так званих фолікулярних клітин, які захищають яйцеклітину та виконують гормоноутворюючу функцію – вони виділяють естрогени.
Фізіологічне положення матки, труб і яєчників забезпечується підвішуючим, фіксуючим та підтримуючим апаратами, що об'єднують очеревину, зв'язки та тканини малого таза. Підвішуючий апарат представлений парними утвореннями, він включає круглу та широку зв'язки матки, власні зв'язки та підвішувальні зв'язки яєчників. Широкі зв'язки матки, власні та підвішувальні зв'язки яєчників утримують матку в середньому положенні. Круглі зв'язки тягнуть дно матки вперед і забезпечують її фізіологічний нахил.
Фіксуючий (якірний) апарат забезпечує положення матки в центрі малого тазу та практично унеможливлює її зміщення в сторони, вперед і назад. Але оскільки зв'язковий апарат відходить від матки в її нижньому відділі, можливі нахили матки в різних напрямках. Фіксуючий апарат включає зв'язки, розташовані в пухкій тканині таза та тягнуться від нижнього відділу матки до бічних, передньої та задньої стінок таза: крижово-магічні, кардинальні, матково-міхурово-міхурово-тазові зв'язки.
Окрім мезоварію, розрізняють такі зв'язки яєчників:
- підвішувальна зв'язка яєчника, раніше позначена як воронко-тазова зв'язка. Це складка очеревини з кровоносними судинами (a. et v. ovarica) та лімфатичними судинами та нервами яєчника, що проходять через неї, натягнута між бічною стінкою таза, поперековою фасцією (в ділянці поділу загальної клубової артерії на зовнішню та внутрішню) та верхнім (трубним) кінцем яєчника;
- Власна зв'язка яєчника проходить між листками широкої маткової зв'язки, ближче до заднього листка, і з'єднує нижній кінець яєчника з латеральним краєм матки. Власна зв'язка яєчника прикріплюється до матки між початком маткової труби та круглою зв'язкою, позаду та над останньою. У товщі зв'язки проходять rr. ovarii, які є кінцевими гілками маткової артерії;
- Апендикулярно-яєчникова зв'язка Зв'язка тягнеться від брижі відростка до правого яєчника або широкої зв'язки матки у вигляді складки очеревини. Зв'язка непостійна і спостерігається у 1/2 - 1/3 жінок.
Опорний апарат представлений м'язами та фасціями тазового дна, що поділяються на нижній, середній та верхній (внутрішній) шари.
Найпотужнішим є верхній (внутрішній) м'язовий шар, представлений парним м'язом, що піднімає анус. Він складається з м'язових пучків, що віялом розходяться від куприка до кісток тазу в трьох напрямках (лобково-куприковий, клубово-куприковий та сіднично-куприковий). Цей шар м'язів також називають тазовою діафрагмою.
Середній шар м'язів розташований між симфізом, лобковою та сідничною кістками. Середній шар м'язів - сечостатева діафрагма - займає передню половину виходу з малого тазу, через яку проходять уретра та піхва. У передньому відділі між її листками розташовані м'язові пучки, що утворюють зовнішній сфінктер уретри, у задньому відділі - м'язові пучки, що йдуть у поперечному напрямку - глибокий поперечний м'яз промежини.
Нижній (зовнішній) шар м'язів тазового дна складається з поверхневих м'язів, форма яких нагадує цифру 8. До них належать бульбокавернозний, сіднично-кавернозний, зовнішній анальний сфінктер та поверхневий поперечний м'яз промежини.
Онтогенез яєчників
Процес росту та атрезії фолікулів починається на 20 тижні вагітності, і до моменту пологів в яєчниках дівчинки залишається до 2 мільйонів ооцитів. До менархе їх кількість зменшується до 300 тисяч. Протягом усього періоду репродуктивного життя не більше 500 фолікулів досягають зрілості та овулюють. Початковий ріст фолікулів не залежить від стимуляції ФСГ, обмежений, і атрезія відбувається швидко. Вважається, що замість стероїдних гормонів основним регулятором росту та атрезії первинних фолікулів є локальні аутокринні/паракринні пептиди. Вважається, що процес росту та атрезії фолікулів не переривається жодними фізіологічними процесами. Цей процес триває в будь-якому віці, включаючи внутрішньоутробний період та менопаузу, і переривається вагітністю, овуляцією та ановуляцією. Механізм, що запускає ріст фолікулів, та їх кількість у кожному конкретному циклі досі незрозумілі.
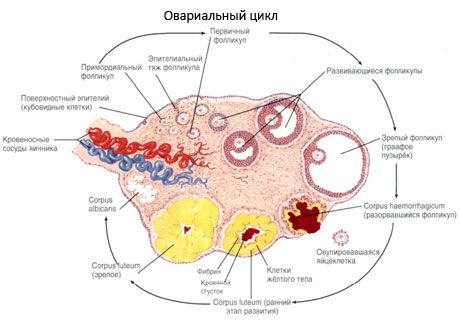
Протягом свого розвитку фолікул проходить кілька стадій розвитку. Первинні статеві клітини походять з ентодерми жовткового мішка, алантоїсу, та мігрують до статевої області ембріона на 5-6 тижні вагітності. В результаті швидкого мітотичного поділу, який триває з 6-8 тижнів до 16-20 тижнів вагітності, в яєчниках ембріона утворюється до 6-7 мільйонів ооцитів, оточених тонким шаром гранульозних клітин.
Преантральний фолікул – ооцит оточений мембраною (Zona pellucida). Гранульозні клітини, що оточують ооцит, починають проліферувати, їх ріст залежить від гонадотропінів і корелює з рівнем естрогенів. Гранульозні клітини є мішенню для ФСГ. На стадії преантрального фолікула гранульозні клітини здатні синтезувати три класи стероїдів: переважно індукує активність ароматази, основного ферменту, що перетворює андрогени на естрадіол. Вважається, що естрадіол здатний збільшувати кількість власних рецепторів, надаючи прямий мітогенний вплив на гранульозні клітини незалежно від ФСГ. Він вважається паракринним фактором, що посилює ефекти ФСГ, включаючи активацію процесів ароматизації.
Рецептори ФСГ з'являються на мембранах гранульозних клітин, як тільки починається ріст фолікула. Зниження або збільшення ФСГ призводить до зміни кількості його рецепторів. Ця дія ФСГ модулюється факторами росту. ФСГ діє через G-білок, аденілатциклазну систему, хоча стероїдогенез у фолікулі переважно регулюється ФСГ, у цьому процесі бере участь багато факторів: іонні канали, рецептори тирозинкінази, фосфоліпазна система вторинних месенджерів.
Роль андрогенів у ранньому розвитку фолікулів є складною. Гранульозні клітини мають андрогенні рецептори. Вони є не тільки субстратом для ароматизації, індукованої ФСГ, в естрогени, але й можуть посилювати ароматизацію при низьких концентраціях. Коли рівень андрогенів підвищується, преантральні гранульозні клітини переважно обирають не шлях ароматизації в естрогени, а простіший шлях андрогенів через 5α-редуктазу для перетворення на андроген, який не може бути перетворений на естроген, тим самим пригнічуючи активність ароматази. Цей процес також пригнічує утворення рецепторів ФСГ та ЛГ, тим самим зупиняючи розвиток фолікулів.
В процесі ароматизації фолікул з високим рівнем андрогенів зазнає процесів атрезії. Ріст і розвиток фолікула залежить від його здатності перетворювати андрогени на естрогени.
За наявності ФСГ домінуючою речовиною фолікулярної рідини будуть естрогени. За відсутності ФСГ – андрогени. ЛГ зазвичай відсутній у фолікулярній рідині до середини циклу. Як тільки рівень ЛГ підвищується, мітотична активність гранульозних клітин знижується, з'являються дегенеративні зміни та підвищується рівень андрогенів у фолікулі. Рівень стероїдів у фолікулярній рідині вищий, ніж у плазмі, і відображає функціональну активність клітин яєчників: гранульозних та тека-клітин. Якщо єдиною мішенню для ФСГ є гранульозні клітини, то ЛГ має багато мішеней – тека-клітини, стромальні та лютеїнові клітини та гранульозні клітини. Як гранульозні, так і тека-клітини мають здатність до стероїдогенезу, але в гранульозних клітинах переважає ароматазна активність.
У відповідь на ЛГ тека-клітини виробляють андрогени, які потім перетворюються гранульозними клітинами на естрогени шляхом ароматизації, індукованої ФСГ.
У міру зростання фолікула тека-клітини починають експресувати гени рецепторів ЛГ, Р450 sec та 3β-гідроксистероїддегідрогенази, інсуліноподібний фактор росту (IGF-1) синергічно з ЛГ збільшує експресію генів, але не стимулює стероїдогенез.
Стероїдогенез в яєчниках завжди залежить від ЛГ. У міру зростання фолікула тека-клітини експресують фермент P450c17, який утворює андроген з холестерину. Гранульозні клітини не мають цього ферменту і залежать від тека-клітин для вироблення естрогенів з андрогенів. На відміну від стероїдогенезу, фолікулогенез залежить від ФСГ. У міру зростання фолікула та підвищення рівня естрогену активується механізм зворотного зв'язку – вироблення ФСГ пригнічується, що, у свою чергу, призводить до зниження активності ароматази фолікула та, зрештою, до атрезії фолікула через апоптоз (запрограмовану загибель клітин).
Механізм зворотного зв'язку естрогенів та ФСГ пригнічує розвиток фолікулів, що почали рости, але не домінантного фолікула. Домінантний фолікул містить більше рецепторів ФСГ, які підтримують проліферацію гранульозних клітин та ароматизацію андрогенів до естрогенів. Крім того, паракринний та аутокринний шляхи виступають важливим координатором розвитку антрального фолікула.
Аутокринний/паракринний регулятор складається з пептидів (інгібін, активін, фолістатин), які синтезуються гранульозними клітинами у відповідь на ФСГ та потрапляють у фолікулярну рідину. Інгібін зменшує секрецію ФСГ; активін стимулює вивільнення ФСГ з гіпофіза та посилює дію ФСГ у яєчнику; фолістатин пригнічує активність ФСГ, можливо, зв'язуючись з активіном. Після овуляції та розвитку жовтого тіла інгібін перебуває під контролем ЛГ.
Ріст і диференціація клітин яєчників залежить від інсуліноподібних факторів росту (IGE). IGF-1 діє на гранульозні клітини, викликаючи збільшення рівня циклічного аденозинмонофосфату (цАМФ), прогестерону, окситоцину, протеоглікану та інгібіну.
IGF-1 діє на тека-клітини, викликаючи збільшення вироблення андрогенів. Тека-клітини, у свою чергу, виробляють фактор некрозу пухлини (TNF) та епідермальний фактор росту (EGF), які також регулюються ФСГ.
EGF стимулює проліферацію гранульозних клітин. IGF-2 є основним фактором росту у фолікулярній рідині, а також у ній містяться IGF-1, TNF-α, TNF-3 та EGF.
Порушення паракринної та/або аутокринної регуляції функції яєчників, ймовірно, відіграє певну роль у порушеннях овуляції та розвитку полікістозу яєчників.
У міру зростання антрального фолікула вміст естрогенів у фолікулярній рідині збільшується. На піку їх збільшення на гранульозних клітинах з'являються рецептори до ЛГ, відбувається лютеїнізація гранульозних клітин і збільшується вироблення прогестерону. Таким чином, у преовуляторному періоді збільшення продукції естрогенів викликає появу рецепторів ЛГ, ЛГ, у свою чергу, викликає лютеїнізацію гранульозних клітин і вироблення прогестерону. Збільшення прогестерону знижує рівень естрогенів, що, очевидно, викликає другий пік ФСГ в середині циклу.
Вважається, що овуляція відбувається через 10-12 годин після піку ЛГ та через 24-36 годин після піку естрадіолу. Вважається, що ЛГ стимулює редукційний поділ ооцита, лютеїнізацію гранульозних клітин та синтез прогестерону та простагландинів у фолікулі.
Прогестерон посилює активність протеолітичних ферментів, які разом з простагландинами беруть участь у розриві стінки фолікула. Індукований прогестероном пік ФСГ дозволяє ооциту вийти з фолікула, перетворюючи плазміноген на протеолітичний фермент плазмін, та забезпечує достатню кількість рецепторів ЛГ для нормального розвитку лютеїнової фази.
Протягом 3 днів після овуляції гранульозні клітини збільшуються в розмірах, і в них з'являються характерні вакуолі, заповнені пігментом - лютеїном. Тека-лютеїнові клітини диференціюються з теки та строми та стають частиною жовтого тіла. Капіляри, що проникають у жовте тіло, розвиваються дуже швидко під впливом факторів ангіогенезу, і з покращенням васкуляризації збільшується вироблення прогестерону та естрогенів. Активність стероїдогенезу та тривалість життя жовтого тіла визначаються рівнем ЛГ. Жовте тіло не є однорідним клітинним утворенням. Окрім 2 типів лютеїнових клітин, воно містить ендотеліальні клітини, макрофаги, фібробласти тощо. Великі лютеїнові клітини продукують пептиди (релаксин, окситоцин) та активніше беруть участь у стероїдогенезі з більшою ароматазною активністю та більшим синтезом прогестерону, ніж малі клітини.
Пік прогестерону припадає на 8-й день після піку ЛГ. Зазначається, що прогестерон та естрадіол секретуються епізодично в лютеїновій фазі у кореляції з пульсуючим виробленням ЛГ. З утворенням жовтого тіла контроль над продукцією інгібіну переходить від ФСГ до ЛГ. Рівень інгібіну збільшується зі збільшенням естрадіолу до піку ЛГ і продовжує зростати після піку ЛГ, хоча рівень естрогену знижується. Хоча інгібін та естрадіол секретуються гранульозними клітинами, вони регулюються різними шляхами. Зниження рівня інгібіну в кінці лютеїнової фази сприяє збільшенню ФСГ протягом наступного циклу.
Жовте тіло зменшується дуже швидко – на 9-11-й день після овуляції.
Механізм дегенерації незрозумілий і не пов'язаний з лютеолітичною роллю естрогенів або з рецепторним механізмом, як це спостерігається в ендометрії. Існує й інше пояснення ролі естрогенів, що виробляються жовтим тілом. Відомо, що естрогени необхідні для синтезу рецепторів прогестерону в ендометрії. Естрогени лютеїнової фази, ймовірно, необхідні для змін, пов'язаних з прогестероном, в ендометрії після овуляції. Недостатній розвиток рецепторів прогестерону, як наслідок недостатнього рівня естрогену, можливо, є додатковим механізмом безпліддя та ранніх втрат вагітності, ще однією формою дефіциту лютеїнової фази. Вважається, що тривалість життя жовтого тіла встановлюється в момент овуляції. І воно, безумовно, регресує, якщо не буде підтримуватися хоріонічним гонадотропіном людини через вагітність. Таким чином, регресія жовтого тіла призводить до зниження рівня естрадіолу, прогестерону та інгібіну. Зниження рівня інгібіну усуває його супресивний вплив на ФСГ; Зниження рівня естрадіолу та прогестерону дозволяє дуже швидко відновити секрецію ГнРГ та усунути механізм зворотного зв'язку з гіпофіза. Зниження рівня інгібіну та естрадіолу разом зі збільшенням ГнРГ призводить до перенасичення ФСГ над ЛГ. Збільшення рівня ФСГ призводить до росту фолікулів з подальшим відбором домінантного фолікула, і починається новий цикл, якщо вагітність не настає. Стероїдні гормони відіграють провідну роль у репродуктивній біології та загальній фізіології. Вони визначають фенотип людини, впливають на серцево-судинну систему, метаболізм кісток, шкіру, загальне самопочуття та відіграють ключову роль у вагітності. Дія стероїдних гормонів відображає внутрішньоклітинні та генетичні механізми, необхідні для передачі позаклітинного сигналу до ядра клітини, щоб викликати фізіологічну реакцію.
Естрогени дифузно проникають через клітинну мембрану та зв'язуються з рецепторами, розташованими в ядрі клітини. Комплекс рецептор-стероїд потім зв'язується з ДНК. У клітинах-мішенях ці взаємодії призводять до експресії генів, синтезу білка та специфічної функції клітин і тканин.