
Вірус імунодефіциту людини (ВІЛ)
Останній перегляд: 23.04.2024

Весь контент iLive перевіряється медичними експертами, щоб забезпечити максимально можливу точність і відповідність фактам.
У нас є строгі правила щодо вибору джерел інформації та ми посилаємося тільки на авторитетні сайти, академічні дослідницькі інститути і, по можливості, доведені медичні дослідження. Зверніть увагу, що цифри в дужках ([1], [2] і т. д.) є інтерактивними посиланнями на такі дослідження.
Якщо ви вважаєте, що який-небудь з наших матеріалів є неточним, застарілим або іншим чином сумнівним, виберіть його і натисніть Ctrl + Enter.
Синдром набутого імунодефіциту був виділений в якості особливого захворювання в 1981 р в США, коли в ряді молодих людей важкі захворювання були викликані мікроорганізмами, непатогенних або слабопатогеннимі для здорових людей. Дослідження імунного статусу хворих виявило у них різке зменшення кількості лімфоцитів взагалі і Т-хелперів особливо. Цей стан отримало назву AIDS (англ. Acquired Immune Deficiency Syndrome - синдром набутого імунодефіциту, або СНІД). Спосіб зараження (статевий контакт, через кров і її препарати) вказував на інфекційний характер захворювання.
Збудник СНІДу був відкритий в 1983 р незалежно один від одного французом Л. Монтаньє, який назвав його LAV Lymphoadenopathy Associated Virus), так як виявив у хворого лимфоаденопатией; і американцем Р. Галло, який назвав вірус HTLV-III (англ. Human T-lymphotropic Virus III - Т-лімфотропний вірус людини III): раніше їм були виявлені лімфотропні віруси I і II.
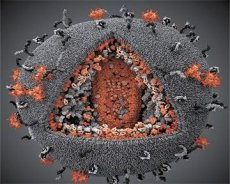
Зіставлення властивостей вірусів LAV і HTLV-III показало їх ідентичність, тому щоб уникнути плутанини вірус отримав в 1986 р назва HIV (англ. Human Immunodeficiency Virus - вірус імунодефіциту людини, або ВІЛ). ВІЛ кулястої форми, його діаметр 110 нм. Оболонка вірусу має форму багатогранника, складеного з 12 п'ятикутників і 20 шестикутників. У центрі і кутах кожного шестикутника розташована молекула глікозильованого протеїну gpl20 (число 120 означає молекулярну масу білка в кілодальтон). Всього на поверхні віріона розташовуються у вигляді своєрідних шипів 72 молекули gpl20, кожна з яких пов'язана з внутрімембранним білком gp41. Ці білки разом з подвійним ліпідним шаром утворюють суперкапсид (мембрану) віріона.
Білки gpl20 і gp41 утворюються в результаті нарізування клітинної протеазой білка-попередника Env. Білок gp41 формує «ніжку» шипа, зв'язуючись цитоплазматическим доменом з розташованим безпосередньо під оболонкою матриксних білком р17МА. Молекули р17, взаємодіючи при дозріванні віріона, утворюють ікосаедр, підстильний оболонку.
У центральній частині віріона білок р24 утворює конусоподібний капсид. Звужена частина капсида за участю білка рб пов'язана з оболонкою віріона. Всередині капсида укладені дві ідентичні молекули вірусної геномної РНК. Вони пов'язані своїми 5'-кінцями з нуклеокапсідний білком p7NC. Цей білок цікавий тим, що має два амінокислотних залишку (мотиву), багатих цистеїном і гистидином і містять атом Zn, - їх називають «цинковими пальцями», так як вони захоплюють молекули геномної РНК для включення в формуються віріони. До складу капсида входять також три ферменту. Ревертаза (RT), або pol-комплекс, включає в себе зворотну транскриптазу, РНК-азу Н і ДНК-залежну ДНК-полімеразу. Ревертаза присутній у вигляді гетеродимера р66 / Р51. Протеаза (PR) - рю, запускає і реалізує процес дозрівання віріона. Інтеграза (IN) - р31, або ендонуклеаза, забезпечує включення провірусної ДНК в геном клітини-господаря. У капсиді міститься також молекула затравочной РНК (тРНКл "3).
РНК-геном в клітині за допомогою зворотної транскриптази перетворюється в ДНК-геном (ДНК-провірус), що складається з 9283 нуклеотидних пар. Він обмежений зліва і справа так званими довгими кінцевими повторами, або LTR (англ. Long terminal repeat): S'-LTR - зліва і З'-LTR - справа. LTR містять по 638 нуклеотидних пар.
Геном ВІЛ складається з 9 генів, частина з яких перекривається кінцями (має кілька рамок зчитування) і має екзонінтронную структуру. Вони контролюють синтез 9 структурних і 6 регуляторних білків.
Значення LTR для вірусного генома полягає в тому, що в них розташовані такі регуляторні елементи, які контролюють його роботу:
- сигнал транскрипції (область промотора);
- сигнал додавання полі-А;
- сигнал кепірованія;
- сигнал інтеграції;
- сигнал позитивної регуляції (TAR для білка ТАТ);
- елемент негативної регуляції (NRE для білка NEF);
- ділянку прикріплення затравочной РНК (тРНК ™ 3) для синтезу мінус-ланцюга ДНК на -кінці; сигнал на 5'-кінці LTR, який служить запалом для синтезу плюс-ланцюга ДНК.
Крім того, в LTR є елементи, які беруть участь в регуляції сплайсингу мРНК, упаковки молекул вРНК в капсид (елемент Psi). Нарешті, при транскрипції генома в довгих мРНК утворюються два сигнали для білка REV, які перемикають синтез білків: CAR - для регуляторних білків і CRS - для структурних білків. Якщо білок REV зв'язується з CAR, синтезуються структурні білки; якщо він відсутній, синтезуються тільки регуляторні білки.
У регуляції роботи генома вірусу особливо важливу роль відіграють такі гени-регулятори і їх білки:
- білок ТАТ, який здійснює позитивний контроль розмноження вірусу і діє через регуляторний ділянку TAR;
- білки NEV і VPU, які здійснюють негативний контроль розмноження через ділянку NRE;
- білок REV, який здійснює позитивно-негативний контроль. Білок REV контролює роботу генів gag, pol, env і здійснює негативну регуляцію сплайсингу.
Таким чином, розмноження ВІЛ знаходиться під потрійним контролем - позитивним, негативним і позитивно-негативним.
Білок VIF визначає інфекційність знову синтезованого вірусу. Він пов'язаний з капсидних білків р24 і присутній в вирионе в кількості 60 молекул. Білок NEF представлений в вирионе невеликим числом молекул (5-10), можливо, пов'язаних з оболонкою.
Білок VPR гальмує клітинний цикл на фазі G2, бере участь в транспорті преінтеграціонних комплексів в ядро клітини, активує деякі вірусні і клітинні гени, підвищує ефективність реплікації вірусу в моноцитах і макрофагах. Місце розташування білків VPR, TAT, REV, VPU в вирионе не встановлено.
Крім власних білків до складу оболонки віріона можуть входити деякі білки клітини-господаря. Білки VPU і VPR беруть участь в регуляції репродукції вірусу.
Антигенні варіанти вірусу імунодефіциту людини (ВІЛ)
Вірус імунодефіциту людини (ВІЛ) дуже мінливий. Навіть з організму одного хворого можуть бути виділені штами вірусу, що істотно розрізняються за антигенними властивостями. Такий мінливості сприяють інтенсивне руйнування клітин CD4 + і потужний антитільний відповідь на ВІЛ-інфекцію. У хворих із Західної Африки виділена нова форма ВІЛ, біологічно близька до ВІЛ-1, але імунологічно відрізняється від нього, - ВІЛ-2. Гомологія первинної структури геномів цих вірусів становить - 42%. ДНК-провірус ВІЛ-2 містить 9671 п. Н., А його LTR - 854 п. Н. ВІЛ-2 згодом виділено і в інших регіонах світу. Перехресного імунітету між ВІЛ-1 і ВІЛ-2 немає. Відомі дві великі форми ВІЛ-1: О (Outlier) і М (Major), останню поділяють на 10 субтипов (AJ). У Росії циркулюють 8 субтипов (А-Н).
 [
[Механізм взаємодії ВІЛ з кліткою
Проникнувши в організм, вірус в першу чергу атакує клітини, що містять специфічний для нього рецептор CD4. Цей рецептор мають у великій кількості Т-хелпери, в меншій - макрофаги і моноцити, особливо до вірусу чутливі Т-хелпери.
Вірус імунодефіциту людини (ВІЛ) розпізнає СD4-рецептори за допомогою свого білка gpl20. Процес взаємодії ВІЛ з кліткою протікає за наступною схемою: рецептор адсорбція -> облямована ямка -> облямований бульбашка -> лизосома. У ній Мембрана віріона зливається з мембраною лізосоми, і нуклеокапсид, звільнений від суперкапсиду, виходить в цитоплазму; на шляху до ядра він руйнується, і вивільняються геномна РНК і асоційовані з нею компоненти серцевини. Далі зворотна транскриптаза синтезує на віріонів РНК мінус-ланцюг ДНК, потім РНК-аза Н руйнує віріони РНК, а вірусна ДНК-полімераза синтезує плюс-ланцюг ДНК. На кінцях ДНК-провируса утворюються 5'-LTR і З'-LTR. ДНК-провірус може перебувати в ядрі деякий час в неактивній формі, але рано чи пізно він за допомогою своєї інтеграли вбудовується в хромосому клітини-мішені. У ній провірус знаходиться в неактивному стані до тих пір, поки даний Т-лімфоцит не буде діяти мікробними антигенами або іншими імунокомпетентними клітинами. Активація транскрипції клітинної ДНК регулюється особливим ядерним фактором (NF-kB). Він є ДНК-зв'язує білком і виробляється у великій кількості при активації і проліферації Т-лімфоцитів і моноцитів. Цей білок зв'язується з певними послідовностями клітинної ДНК і подібними послідовностями LTR ДНК-провируса і індукує транскрипцію як клітинної ДНК, так і ДНК-провируса. Індукуючи транскрипцію ДНК-провируса, він і здійснює перехід вірусу з неактивного стану в активне і відповідно персистентной інфекції - в продуктивну. Перебування провируса в неактивному стані може тривати дуже довго. Активація вірусу є критичним моментом в його взаємодії з клітиною.
З моменту проникнення вірусу в клітину починається період ВІЛ-інфекції - вирусоносительства, яке може тривати 10 і більше років; а з моменту активації вірусу починається хвороба - СНІД. За допомогою своїх регуляторних генів і їх продуктів вірус починає активно розмножуватися. ТАТ-білок може підвищити швидкість репродукції вірусу в 1000 разів. Транскрипція вірусу має складний характер. Вона включає утворення як повнорозмірних, так і субгеномних мРНК, сплайсинг мРНК, а далі відбувається синтез структурних і регуляторних білків.
Синтез структурних білків відбувається так. Спочатку синтезується поліпротеїнів-попередник Pr55Gag (білок з м. М. 55 кД). Він містить 4 основних домену: матриксний (МА), капсидний (СА), нуклеокапсідний (NC) і домен рб, з яких в результаті нарізування Pr55Gag вірусної протеази (вона самовирезается з іншого білка-попередника - Gag-Pol) утворюються відповідно структурні білки р17 , р24, р7 і рб. Утворення поліпротеїну Pr55Gag - головна умова формування вірусних частинок. Саме цей білок визначає програму морфогенезу віріона. Вона включає послідовно стадії транспорту поліпротеїну Gag до плазматичної мембрани, взаємодії з нею і білок-білкових взаємодій при формуванні вірусної частинки і її брунькування. Pr55Gag синтезується на вільних полірібосомамі; молекули білка транспортуються до мембрани, на якій заякорюють своїми гідрофобними ділянками. Основну роль у створенні нативной конформації Gag-білка грає СА-домен. NC-домен забезпечує включення (за допомогою своїх «цинкових пальців») 2 молекул геномної РНК до складу формується вірусної частинки. Молекула поліпротеїну спочатку димеризуется завдяки взаємодії матриксних доменів. Потім димери об'єднуються в гексамерние (з 6 одиниць) комплекси в результаті взаємодії доменів СА і NC. Нарешті, гексамери, з'єднуючись бічними поверхнями, утворюють незрілі віріони сферичної форми, всередині яких міститься геномна вірусна РНК, захоплена NC-доменом.
Інший білок-попередник Prl60Gag-Pol (білок з м. М. 160 кБ) синтезується в результаті зсуву рамки зчитування рибосомою при трансляції -кінця гена gag в області, розташованої безпосередньо перед ділянкою, що кодує білок рб. Цей поліпротеїнів Gag-Pol містить неповну послідовність Gag-білка (1 - 423 амінокислоти) і послідовності Pol, які включають домени PR, RT і IN. Молекули поліпротеїну Gag-Pol також синтезуються на вільних полірібосомамі і транспортуються до плазматичної мембрани. Поліпротеїнів Prl60Gagpol містить всі властиві поліпротеїнів Gag сайти міжмолекулярних взаємодій і сайти зв'язування з мембраною. Тому молекули поліпротеїну Gag-Pol зливаються з мембраною і поряд з Gag-молекулами включаються в формуються віріони, в результаті чого з'являється активна протеаза і починається процес дозрівання віріона. Протеаза ВІЛ-1 високоактивних тільки у вигляді димеру, тому для її самовирезанія з Prl60Gag-Pol потрібно димеризація цих молекул. Дозрівання віріона полягає в тому, що звільнилася активна протеаза розрізає prl60Gag-Pol і Gag55 в впізнаваних нею сайтах; утворюються білки р17, р24, р7, Р6, ревертаза, інтеграли і відбувається їх асоціація в вірусну структуру.
Білок Env синтезується на рибосомах, пов'язаних з мембранами ендоплазматичної, потім він глікозіліруется, розрізається клітинної протеазой на gp120 і gp41 і транспортується на клітинну поверхню. При цьому gp41 пронизує мембрану і зв'язується матриксного доменами молекули Gag-білка, асоційованими з внутрішньою поверхнею мембрани. Цей зв'язок зберігається і в зрілому вирионе.
Таким чином, складання вірусних частинок полягає в агрегації білків-попередників і пов'язаних з ними молекул РНК на плазматичній мембрані клітини-господаря, утворенні незрілих віріонів і їх вивільненні шляхом брунькування з клітинної поверхні. При брунькування вирион оточує себе клітинної мембраною, в яку вбудовані молекули gp41 і gp120. Під час брунькування або, можливо, після вивільнення віріонів відбувається їх дозрівання, яке здійснюється за допомогою вірусної протеази і полягає в протеолітичних нарізанні білків-попередників Pr55Gag і Prl60Gag-Pol на білки зрілого вірусу і їх асоціації в певні структурні комплекси. Провідну роль в процесах морфогенезу вірусу грає поліпротеїнів-попередник Pr55Gag, який організовує і здійснює збірку незрілого віріона; процес його дозрівання завершує специфічна вірусна протеаза.
Причини імунодефіциту
Однією з основних причин імунодефіциту при ВІЛ-інфекції є масова загибель Т-хелперів. Вона настає внаслідок наступних подій. По-перше, заражені вірусом Т-хелпери гинуть внаслідок апоптозу. Вважається, що у хворих на СНІД реплікація вірусу, апоптоз і зниження числа Т-хелперів пов'язані між собою. По-друге, Т-кілери розпізнають і руйнують Т-клітини, інфіковані вірусом або несуть на собі адсорбовані молекули gpl20, а також вірусінфіцірованние і не заражені вірусом Т-хелпери, які утворюють симпласти (синцитій), що складаються з декількох десятків клітин (частина з них гине в результаті розмноження в них вірусів). Внаслідок руйнування великої кількості Т-хелперів відбувається зниження експресії мембранних рецепторів у В-лімфоцитів до інтерлейкіну-2, порушується синтез різних інтерлейкінів (факторів росту і диференціювання В-лімфоцитів - IL-4, IL-5, IL-6 і ін.), в результаті чого порушується функція системи Т-кілерів. Відбувається придушення активності систем комплементу і макрофагів. Інфіковані вірусом макрофаги і моноцити довго не гинуть, але вони не здатні видаляти вірус з організму. Нарешті, через структурного і антигенного подібності gpl20 з рецепторами деяких епітеліальних клітин організму (в тому числі з рецепторами трофобластов, які опосередковують трансплантаційну передачу ВІЛ) відбувається синтез антірецепторних антитіл з широким спектром дії. Такі антитіла здатні блокувати різні клітинні рецептори і ускладнюють перебіг хвороби аутоімунними розладами. Наслідком ВІЛ-інфекції є ураження всіх основних ланок системи імунітету. Такі хворі стають беззахисними проти самих різних мікроорганізмів. Це призводить до розвитку у них опортуністичних інфекцій і пухлинних захворювань. Для хворих на ВІЛ-інфекцією підвищений ризик розвитку раку щонайменше трьох типів: саркоми Капоші; карциноми (включаючи рак шкіри); По-клітинної лімфоми, що виникає через злоякісного переродження В-лімфоцитів. Однак ВІЛ володіє не тільки лімфоціто-, але і нейротропностью. Він проникає в клітини центральної нервової системи (астроцити) як шляхом рецепторопосредованного ендоцитозу, так і при фагоцитозі астроцитами вірусінфіцірованних лимфобластов. При взаємодії вірусу з астроцитами також утворюються симпласти, що сприяють поширенню збудника по міжклітинних каналам. У макрофагах і моноцитах вірус може зберігатися тривалий час, тому вони служать резервуаром і розповсюджувачами його в організмі, будучи здатні проникати в усі тканини. Інфікованим макрофагів належить головна роль в заметі ВІЛ в ЦНС і її ураження. У 10% хворих первинні клінічні синдроми пов'язані з ураженням ЦНС і проявляються у вигляді деменції (недоумства). Таким чином, для людей, уражених ВІЛ-інфекцією, характерні 3 групи захворювань - опортуністичні інфекції, пухлинні хвороби і ураження ЦНС.
Епідеміологія ВІЛ-інфекції
Джерелом ВІЛ-інфекції є тільки людина - хворий або вірусоносій. Вірус імунодефіциту людини (ВІЛ) міститься в крові, спермі, цервікальної рідини; у годуючих матерів - в грудному молоці. Зараження відбувається статевим шляхом, через кров і її препарати, а також від матері до дитини до пологів, під час і після пологів. Випадки зараження вірусом через харчові продукти, напої та через укуси комах не відомі.
Поширенню СНІДу сприяє наркоманія. Зараженість ВІЛ зростає з кожним роком. За даними ВООЗ, з 1980 по 2000 р 58 млн осіб було інфіковано ВІЛ. Тільки протягом 2000 року в світі було інфіковано 5,3 млн, а померли від СНІДу 3 млн чоловік. У Росії на 1 січня 2004 року було зареєстровано 264 тис. ВІЛ-інфікованих людей. Половина осіб, заражених ВІЛ, помирає протягом 11-12 років з моменту зараження. На початку 2004 р з кожних 100 тис. Громадян Росії близько 180 жили з діагнозом «ВІЛ-інфекція». Прогнозується, що при такому рівні захворюваності сумарне число ВІЛ-інфікованих в Росії до 2012 р складе 2,5-3 млн осіб. Складність боротьби з ВІЛ-інфекцією залежить від ряду причин: по-перше, ще немає ефективних методів її лікування і специфічної профілактики; по-друге, інкубаційний період при ВІЛ-інфекції може перевищувати 10 років. Його тривалість залежить від моменту активації Т-лімфоцити і міститься в його хромосомі ДНК-провируса. Поки неясно, приречений чи кожен інфікований вірусом на СНІД або можливо тривале вірусоносійство без захворювання (що здається малоймовірним). Нарешті, існує кілька вірусів імунодефіциту людини (ВІЛ-1, ВІЛ-2), антигенні відмінності між якими запобігають формуванню перехресного імунітету. Виявлення вірусу імунодефіциту мавп (ВІМ) пролило світло на питання про походження ВІЛ. ВІО по організації генома схожий з ВІЛ, але істотно відрізняється по нуклеотидної послідовності. ВІЛ-2 по серологічним властивостям займає проміжне положення між ВІЛ-1 і ВІО, а по нуклеотидної послідовності виявився ближче до ВІО. У зв'язку з цим В. М. Жданов припустив, що віруси ВІЛ-1, ВІЛ-2 і ВІО походять від спільного предка. Не виключено, на думку Р. Галло, що один з ВІО якимось чином потрапив в організм людини, де сталося чимало мутацій, в результаті яких виникли ВІЛ-1, ВІЛ-2 і інші його форми.
Симптоми ВІЛ-інфекції
Вірусу імунодефіциту людини властиві деякі особливості, від яких багато в чому залежить патогенез захворювання. Вірус має дуже високою швидкістю розмноження, яка визначається його регуляторними елементами (за 5 хв в активній стадії синтезується до 5000 віріонів). Завдяки наявності білка злиття (gp41) вірус індукує утворення великих синцитіальних структур за рахунок злиття інфікованих і неінфікованих Т-хелперів, наслідком чого є їх масова загибель. Утворені в великій кількості молекули білка gpl20 вільно циркулюють в крові і зв'язуються з рецепторами неінфікованих Т-хелперів, в результаті чого вони також розпізнаються і знищуються Т-кілерами. Вірус може поширюватися по міжклітинних каналах з клітки в клітку, в цьому випадку він стає мало доступний антитіл.
Клінічні критерії ВІЛ-інфекції
У дорослих ВІЛ-інфекцію встановлюють при наявності у них щонайменше двох серйозних симптомів у поєднанні хоча б з одним незначним симптомом і при відсутності інших відомих причин імунодефіциту (рак, вроджений імунодефіцит, важка форма голодування і т. П.). До серйозних симптомів відносять:
- схуднення на 10% і більше;
- тривалий гарячковий стан, що чергується або постійне;
- хронічна діарея.
Незначні симптоми: завзятий кашель, генералізований дерматит, рецидивний оперізувальний герпес, кандидоз ротової порожнини і глотки, хронічний простий герпес, генералізована лімфаденопатія. Діагноз СНІДу ставлять при наявності однієї лише саркоми Капоші, криптококового менінгіту, пневмоцістноі пневмонії. На клінічну картину хвороби впливає приєдналася опортуністична інфекція.
Методи культивування вірусу імунодефіциту людини (ВІЛ)
ВІЛ-1 і ВІЛ-2 вдається культивувати в клітинах тільки одного клону ТСБ4-лімфоцитів - Н9, отриманого з лейкозних ТСВ4-лімфоцитів. Для цих же цілей можуть бути використані і моношарова культури клітин астроцитів, в яких ВІЛ-1 добре розмножується. З тварин до ВІЛ-1 сприйнятливі шимпанзе.
Резистентність вірусу у зовнішньому середовищі невелика. Він гине під впливом сонячних променів і УФ-опромінення, руйнується при 80 ° С протягом 30 хв, при обробці звичайно застосовуються дезінфікуючими речовинами - протягом 20-30 хв. Для знезараження віруссодержащего матеріалу необхідно користуватися мікобактеріціднимі дезінфікуючими речовинами, оскільки вони ефективні проти мікроорганізмів, що володіють найвищою резистентністю.
Лабораторна діагностика ВІЛ-інфекції
Основним способом діагностики вірусоносійства і ВІЛ-інфекції є імуноферментний метод. Однак у зв'язку з тим, що gpl20 має структурний і антигенну схожість з рецепторами деяких клітин людини, в тому числі з рецепторами, які здійснюють транспорт імуноглобулінів через епітеліальні клітини слизових оболонок, в організмі можуть з'являтися антитіла, споріднені антитіл проти gpl20. В цьому випадку можуть бути хибнопозитивних результати ІФМ. Тому все позитивно реагують сироватки досліджуваних піддаються додатковому аналізу за допомогою методу иммуноблотинга, або вестернблотінга. В основі цього методу лежить ідентифікація досліджуваних антитіл після електрофоретичного розділення їх і подальшого тестування за допомогою мічених антивидових антитіл. Вірусологічні методи мало застосовується через складність культивування вірусу. Клон лімфоцитів Н9 використовується для отримання вірусних антигенів - необхідних компонентів діагностичних тест-систем. Метод ЦПР дозволяє виявити вірус вже на ранньому етапі вирусемии.
Лікування ВІЛ-інфекції
Необхідно знайти або синтезувати препарати, ефективно пригнічують активність зворотної транскриптази (ревертази) або вірусної протеази. Вони запобігали б утворення ДНК-провируса і (або) інгібували внутрішньоклітинний розмноження вірусу. Сучасна стратегія лікування ВІЛ-інфікованих заснована на принципі комбінованого застосування препаратів, що пригнічують вірусну протеазу (один з препаратів) і ревергазу (2 різних препарату), - комбінована (потрійна) терапія. У Росії для лікування ВІЛ-інфікованих рекомендовано одночасне застосування 2 вітчизняних препаратів: Фосфазід і Кріксіван, специфічно пригнічують репродукцію ВІЛ на ранніх і пізніх стадіях розмноження, особливо при зниженій активності азидотимидина.
Проблема специфічної профілактики полягає в необхідності створення вакцини, яка б забезпечувала формування ефективного клітинно-опосередкованого імунітету на основі вирусспецифических цитотоксичних лімфоцитів без скільки-небудь істотної продукції антитіл. Такий імунітет забезпечують Thl-хелпери. Можливо, що антитіла, в тому числі і вируснейтрализующие, не тільки не ефективні в придушенні ВІЛ-інфекції, але при високому рівні пригнічують клітинно-опосередкований імунітет. Тому анти-ВІЛ-вакцина повинна відповідати перш за все двом основним вимогам: а) бути абсолютно безпечною і б) стимулювати активність Т-цитотоксичних лімфоцитів. Вивчається ефективність різних варіантів вакцин, отриманих з убитих (інактивованих) вірусів і з окремих антигенів, з високими протектівнимі властивостями. Такі антигени можуть бути або виділені з самих віріонів, або синтезовані хімічно. Запропоновано вакцина, створена на основі методів генної інженерії. Вона являє собою рекомбінантний вірус осповакціни, що несе гени ВІЛ, відповідальні за синтез антигенів з сильними імуногенні властивості. Вирішення питання про ефективність цих вакцин вимагає значного часу через велику тривалості інкубаційного періоду ВІЛ-інфекції та високу мінливість збудника. Створення високоефективної вакцини проти ВІЛ - невідкладна фундаментальна проблема.